
Motivational-Emotional Pathway
In (Vrtička et al., 2017) the authors propose a motivational-emotional pathway to compassion described by the movement through disparate states:
- mimicry
- emotional contagion
- empathy
- compassion
Mimicry refers to the tendency for individuals to physically mimic the other person’s expression when they are displaying a certain emotion. This mimicry allows individuals to more easily feel the specific emotion that the other is displaying. For instance, a friend may be opening up about a sad event and a natural response is for the facial features of the listener to tend towards a sad expression.
The actual experiencing of sadness (or any other emotion) by the listener is then referred to as emotional contagion. This allows the listener to vicariously share the emotion that the other is experiencing. It is important to note that this is not only for negative emotions but also for positive emotions. When we are around someone who is particularly happy it often has the effect of making us smile—mimicry—followed by feeling a similar state of happiness.
Empathy is when we feel the emotion of the other person while simultaneously knowing that such a feeling is stemming from the other, i.e. we do not mistake the sadness that is arising in us as being directly due to our own situation but indirectly due to the other persons.
Compassion can arise if we then choose to be fully present to this other person’s suffering with a sincere wish to alleviate it, stemming from a place of kindness and warmth. The benefit of this response is that it is actually rewarding to do so.
The alternative to this is empathic distress. This is when the feeling of the emotion stemming from the other person begins to overwhelm us and it feels like we are the one who is suffering. We mistake the other person’s state for our own. The effect of this can often lead to avoidance and resistance to the other person’s situation. Rather than engaging with it and opening ourselves up, we close ourselves off and avoid it. Although this may feel safer or more manageable in the short term, it tends to be less rewarding in the long term, as we are shutting ourselves off from the capacity to be present with another person and forgoing the opportunity to alleviate the other person’s suffering.

fMRI Meta-analysis
There have been a variety of fMRI studies on compassion that have taken place. A recent meta-analysis (Kim et al., 2020) selected 16 of these studies under their selection process to figure out what brain regions are most commonly activated during compassion states. They found seven broad regions to be most commonly activated during these studies:
- middle frontal gyrus
- inferior frontal gyrus
- medial frontal gyrus
- anterior cingulate cortex
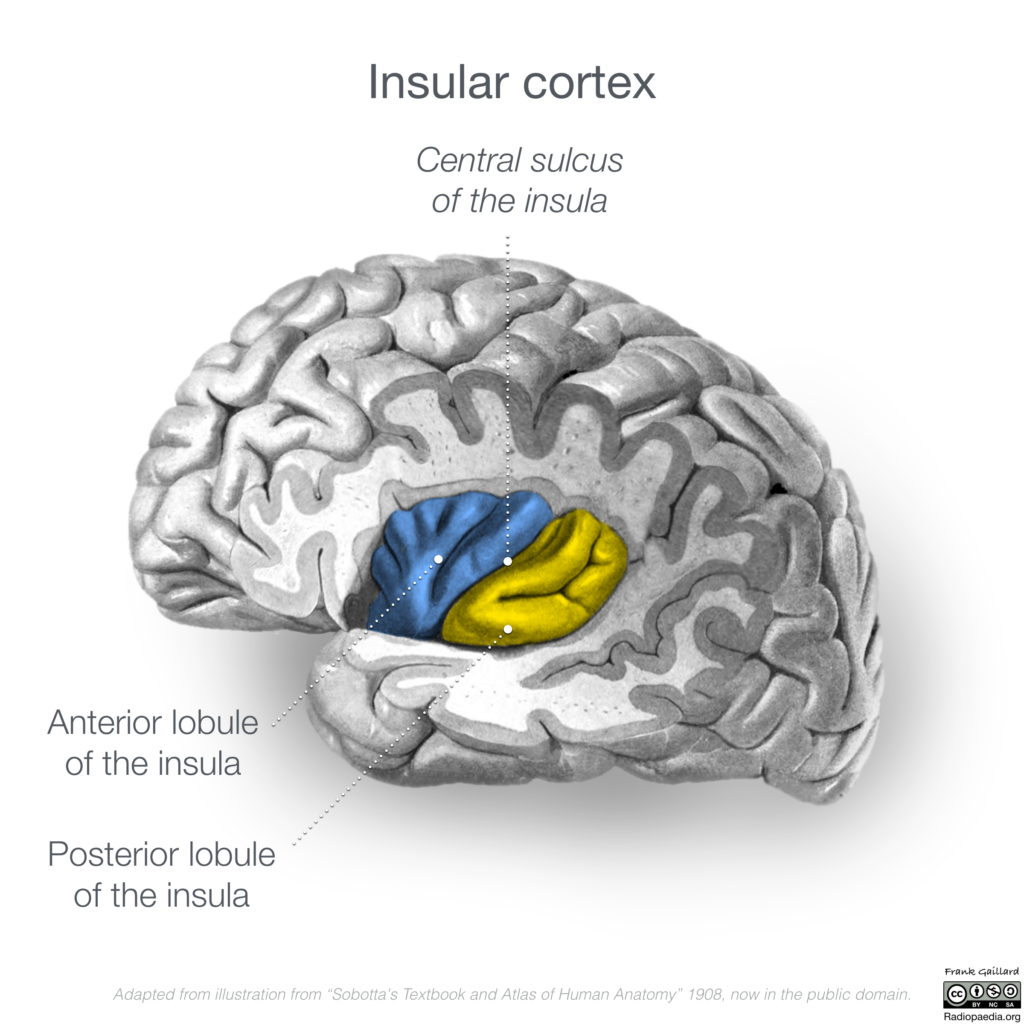
- anterior insular cortex
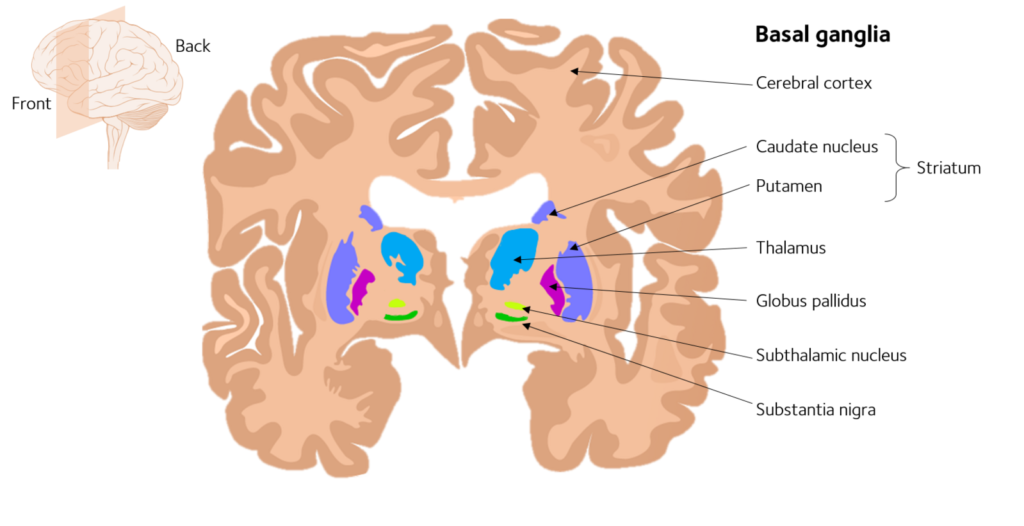
- basal ganglia/thalamus circuitry
- other
The highest peaks of activation were found in:
- periaqueductual grey (part of the basal ganglia/thalamic circuitry)
- anterior insular cortex
- anterior cingulate cortex
- inferior frontal gyrus.
And lastly, they found that a few regions often mentioned in relation to compassion were not found to be amongst the most commonly activated:
- dorsolateral prefrontal cortex (DLPC)
- orbito frontal cortex (OFC)
- amygdala
The authors propose that this latter finding may be related to the difference between participants while engaging in loving-kindness or compassion practices during studies.
We will briefly review the understanding of why each of these brain regions may be part of the neural system for compassion.
Anterior Cingulate Cortex and Anterior Insular Cortex
The regions of the anterior cingulate cortex and anterior insular cortex are nodes in the salience network, a network which is often active when investigating the neural responses to experienced and observed pain. It is possible that this network is commonly active due to the experimental setup; many experiments make use of negative stimuli, e.g. sad faces, to evoke feelings of compassion within participants. So, this processing of affective, emotional stimuli, may activate the network rather than the compassionate state itself. However, there is evidence from experimental paradigms that do not utilise such negative stimuli that this network is still active, so, the salience network may be active during compassion due to the processing of negative emotion—in this case the suffering of another person.

Frontal Gyri (middle, inferior, and medial)
The frontal gyri are associated with a range of functions that include language, attention, and the capacity to monitor emotion and valence (Kim et al., 2020). It is not yet clear what the overall functional role of these regions are in compassion. There is some evidence provided in human lesion models (Shdo et al., 2018) for a relationship between the inferior frontal gyrus and prosocial motivation, a concept similar to compassion.






Middle column: Inferior frontal gyrus dorsal view (above) and lateral view (below)
Right column: Medial frontal gyrus medial view.
Basal Ganglia and Thalamus
The basal ganglia (or basal nuclei) is a cluster of subcortical nuclei composed of:
- caudate nucleus (part of the dorsal striatum)
- putamen (part of the dorsal striatum)
- globus pallidus
- subthalamic nucleus
- substantial nigra
- nucleus accumbens (part of the ventral striatum, sometimes considered to not be part of the basal ganglia and instead part of the broader limbic system.)
- olfactory tubercle (part of the ventral striatum, sometimes considered to not be part of the basal ganglia and instead part of the broader limbic system.)
It is normally associated with reward, cognition, and motor control.
The thalamus is a separate structure in the forebrain that is often referred to as a ‘gateway’ to the cerebral cortex because incoming sensory information from the eyes, ears, and skin all relay through the thalamus before being transmitted to the cortex (Bear et al., 2020).
The two regions form an overlapping network called the cortico-basal ganglia-thalamus-cortico (CBGTC) loop.
According to (Kim et al., 2018) dysfunction of the basal ganglia and/or thalamus “been associated with deficits in emotional expression and processing, the inability to associate emotion with behaviour (Levy and Dubois, 2006), and the inability to detect salient emotional cues (Paulmann et al., 2013). Furthermore, convergent evidence from stroke-patient models has established basal ganglia impairment is associated with significantly greater affective (i.e., apathy, depression) and cognitive deficits than controls (Onoda et al., 2010; Paradiso et al., 2013).”
Further research may allow elucidation of the exact contribution that each of these subregions has for compassion.
Midbrain and Periaqueductal Grey
The periaqueductal grey is a column of grey matter that surrounds the cerebral aqueduct of Sylvius in the midbrain (Hattingh et al., 2024) and receives input from several brain structures, many of them appropriate for transmitting signals related to emotional status, and is important in the control of the somatic pain sensations (Bear et al., 2020).
Interestingly, this region formed the largest of the foci (areas of activation) in the meta-analysis but was only activated in four of the studies. It may be that the midbrain and periaqueductal grey are not necessary for compassion but act to enhance it as these regions are implicated in underlying mammalian care systems.
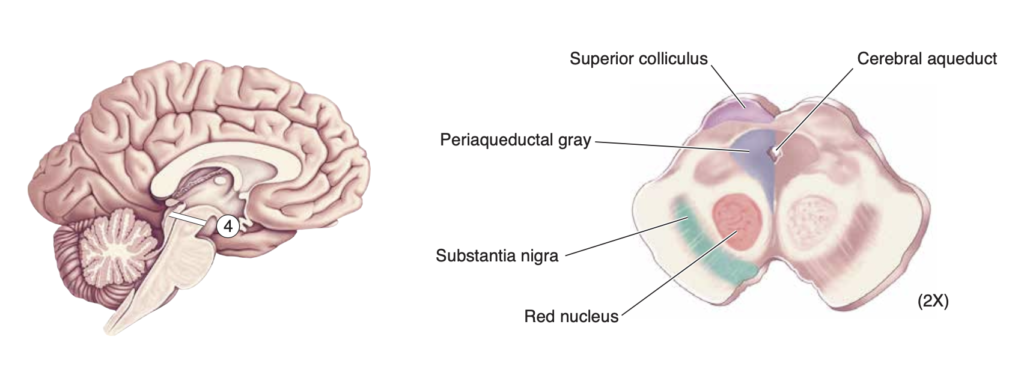
Right: Periaqueductal grey surrounding the cerebral aqueduct and other related structures.
fMRI Paradigms
The authors (Kim et al., 2020) note that there are multiple experimental paradigms for researching compassion in use with fMRI. Based on their analysis there are four different operationalizations of compassion occurring within the context of these studies: stimulus, mindset, target, and multimodal.
fMRI Experimental Setups
It is worth understanding the experimental paradigm that most of these studies are using. In general, they consist of a stimulus (e.g. images, videos, written descriptions) that depicts someone suffering, a contrast stimulus that depicts something different than this, i.e. a neural or positive stimulus, and either an instruction on how to engage with these stimuli (e.g. engage in compassion) or no instruction at all. Below, there are diagrams depicting this general structure of experimental setup.
Researchers are then in possession of fMRI data when participants are witnessing suffering and fMRI data when participants are not witnessing suffering. Often the participants are asked to report on the level of compassion they experience during presentation of different stimuli. Researchers can then compare these two datasets in order to gain an understanding of what brain regions appear to be active during the witnessing of another person’s suffering compared to witnessing non-suffering/normal situations. The idea is that the suffering will give rise to compassion (whether requested or not), and we will have data showing the brain activity related to this.

Experimental Challenges
It is important to distinguish between different responses to suffering and attempt to isolate the state of compassion. As is noted above in 1 Motivational-emotional pathway, one can also have aversive responses to suffering as empathic distress, people may get angry, perhaps experience sorrow, etc. Therefore, in most experiments the participants are normally either instructed to respond compassionately and/or notified that they will report on their levels of compassion which may lead them to generate it, or only asked afterwards. The issue with all of this is that unless the participants gathered are known to be particularly proficient at engaging in compassion then we may not be measuring compassion at all, but a host of other things. In some studies, participants are recruited who are adept at being this state so as to ensure that we are definitely gathering data on it.
References
Bear, M., Connors, B., & Paradiso, M. A. (2020). Neuroscience: exploring the brain, enhanced edition: exploring the brain. Jones & Bartlett Learning.
Beauregard, M., Courtemanche, J., Paquette, V., & St-Pierre, É. L. (2009). The neural basis of unconditional love. Psychiatry Research: Neuroimaging, 172(2), 93–98. https://doi.org/10.1016/j.pscychresns.2008.11.003
Engen, H. G., & Singer, T. (2015). Compassion-based emotion regulation up-regulates experienced positive affect and associated neural networks. Social Cognitive and Affective Neuroscience, 10(9), 1291–1301. https://doi.org/10.1093/scan/nsv008
FeldmanHall, O., Dalgleish, T., Evans, D., & Mobbs, D. (2015). Empathic concern drives costly altruism. Neuroimage, 105, 347–356. https://doi.org/10.1016/j.neuroimage.2014.10.043
Hattingh C, Hacking C, Al Kabbani A, et al. Periaqueductal grey matter. Reference article, Radiopaedia.org (Accessed on 27 Feb 2024) https://doi.org/10.53347/rID-34472
Kim, J. J., Cunnington, R., & Kirby, J. N. (2020). The neurophysiological basis of compassion: An fMRI meta-analysis of compassion and its related neural processes. Neuroscience & Biobehavioral Reviews, 108, 112-123.
Kim, J.-W., Kim, S.-E., Kim, J.-J., Jeong, B., Park, C.-H., Son, A. R., Song, J. E., & Ki, S. W. (2009). Compassionate attitude towards others’ suffering activates the mesolimbic neural system. Neuropsychologia, 47(10), 2073–2081. https://doi.org/10.1016/j.neuropsychologia.2009.03.017
Klimecki, O. M., Leiberg, S., Lamm, C., & Singer, T. (2013). Functional Neural Plasticity and Associated Changes in Positive Affect After Compassion Training. Cerebral Cortex, 23(7), 1552–1561. https://doi.org/10.1093/cercor/bhs142
Klimecki, O. M., Leiberg, S., Ricard, M., & Singer, T. (2014). Differential pattern of functional brain plasticity after compassion and empathy training. Social Cognitive and Affective Neuroscience, 9(6), 873–879. https://doi.org/10.1093/scan/nst060
Levy, R., & Dubois, B. (2006). Apathy and the functional anatomy of the prefrontal cortex–basal ganglia circuits. Cerebral cortex, 16(7), 916-928.
Lutz, A., Brefczynski-Lewis, J., Johnstone, T., & Davidson, R. J. (2008). Regulation of the Neural Circuitry of Emotion by Compassion Meditation: Effects of Meditative Expertise. PLOS ONE, 3(3), e1897. https://doi.org/10.1371/journal.pone.0001897
Novak, L., Malinakova, K., Mikoska, P., van Dijk, J. P., & Tavel, P. (2022). Neural correlates of compassion–An integrative systematic review. International Journal of Psychophysiology, 172, 46-59.
Onoda, K., Kuroda, Y., Yamamoto, Y., Abe, S., Oguro, H., Nagai, A., … & Yamaguchi, S. (2010). Post-stroke apathy and hypoperfusion in basal ganglia: SPECT study. Cerebrovascular Diseases, 31(1), 6-11.
Paradiso, S., Ostedgaard, K., Vaidya, J., Ponto, L. B., & Robinson, R. (2013). Emotional blunting following left basal ganglia stroke: the role of depression and fronto-limbic functional alterations. Psychiatry Research: Neuroimaging, 211(2), 148-159.
Paulmann, S., Bleichner, M., & Kotz, S. A. (2013). Valence, arousal, and task effects in emotional prosody processing. Frontiers in psychology, 4, 345.
Shdo, S. M., Ranasinghe, K. G., Gola, K. A., Mielke, C. J., Sukhanov, P. V., Miller, B. L., & Rankin, K. P. (2018). Deconstructing empathy: neuroanatomical dissociations between affect sharing and prosocial motivation using a patient lesion model. Neuropsychologia, 116, 126-135.
Vrtička, P., Favre, P., & Singer, T. (2017). Compassion and the brain. Compassion: Concepts, research and applications, 135-150.
Weng, H. Y., Fox, A. S., Shackman, A. J., Stodola, D. E., Caldwell, J. Z. K., Olson, M. C., Rogers, G. M., & Davidson, R. J. (2013). Compassion training alters altruism and neural responses to suffering. Psychological Science, 24(7), 1171–1180. https://doi.org/10.1177/0956797612469537